





.![]()

Augalas grobuonis Plumbago auriculata turi liaukas, galinčias užmušti smulkius organizmus – paprastai smulkius vabzdžius, prisiartinančius per daug arti žiedų. Jis gali išskirti virškinamuosius fermentus, bet ne taip, kaip tai daro tikrieji vabzdžiaėdžiai (Stoltzfus et al. 2002 PDF). Plumbago artimas kai kurioms vabzdžiaėdžių gentims, įskaitant saulašares.
Didžioji dauguma vabzdžiaėdžių augalų rūšių yra Gvazdikiečių (Caryophyllales) eilėje. DNR tyrimais nustatyta artima giminystė tarp gvazdikiečių eilės vabzdžiaėdžių augalų ir tokių aukštą dirvožemio druskingumą toleruojančių šeimų, kaip Frankeniaceae (frankenijiniai), Tamaricaceae (eglūniniai), Plumbaginaceae (pliumbaginiai) ir Polygonaceae (rūgtiniai) atstovų, o taip pat kai kurių negausių augalų šeimų. Didelį druskingumą toleruojančios augalų rūšys gali augti labai druskinguose dirvožemiuose prie vandenynų ar sūrokų vandens telkinių, išskirdami druskų perteklių pro lapus.
Analizuojant DNR tyrimų rezultatus, sunku nustatyti, kuri druskingumą toleruojanti šeima yra artimiausia gvazdikiečių vabzdžiaėdžiams. Jų atsiskyrimas prasidėjo per daug senai, kad būtų įmanoma tai tiksliai nuspėti. Pliumbaginių šeima šiuo atveju dažniausiai analizuojama, nes joje yra daug žinomų sodo augalų, tokių kaip Plumbago auriculata, bei daug rūšių, turinčių panašius su vabzdžiaėdžių šeimomis požymius.
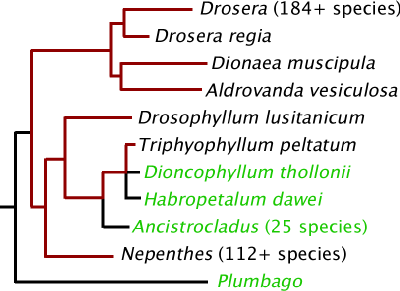
DNR kladograma su juodai pažymėtomis vabzdžiaėdžių gvazdikiečių linijomis ir raudonomis ne vabzdžiaėdžių linijomis. Linijų ilgis proporcingas genetiniams pokyčiams.
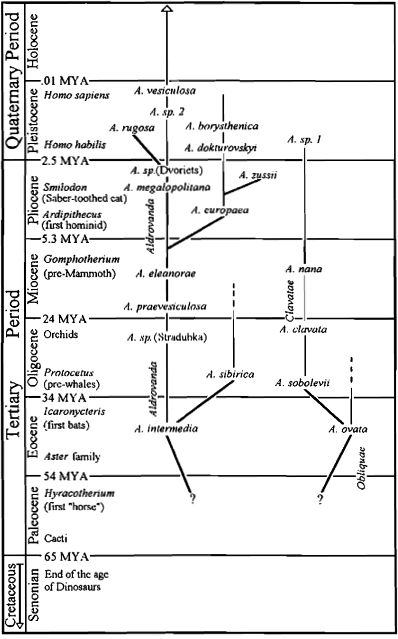
Aldrūnės evoliucija (paveikslėlį galima padidinti). Originali Barry Rice ir Jan Schlauer schema, paremta Degreef (1997 PDF) straipsniu. Iškasenos, priskiriamos Paleoaldrovanda splendens, dabar kaip manoma yra iškastiniai vabzdžių kiaušiniai (Hermanova ir Kvacek 2010). Ši „rūšis” iš schemos pašalinta.
Ką ši kladograma parodo? Augalai su gleivių pagrindu veikiančiomis gaudyklėmis yra artimi aktyvias spąstų gaudykles turintiems, o išskiriantys lipnų skystį artimesni ąsočiais gaudantiems aukas – genčiai, bent dalį savo gyvenimo augančiai kaip lianos (prisiminkite Tarzaną, keliaujantį per džiungles), o nevabzdžiaėdės lianos su vabzdžiaėdžiais galimai turi bendrą protėvį. Kada šis bendras protėvis augo, kaip atrodė, ir kaip iš jo išsivystė tai, ką dabar turime?
Juda pasaulis, juda augalai
Judant žemynams, atskirose jų dalyse keičiasi ir klimatas. Norėdami neišnykti, augalai turi prisitaikyti. Dalinai dėl šios priežasties reliktinės rūšys dažnai sutinkamos kalnuose. Yra paprasčiau augant judėti kalno šlaitu viršun ar apačion, nei nukeliauti šimtus, jei ne tūkstančius, kilometrų į šiaurę ar pietus. Kartais augalai „pabėga” iš kalnų ar žemynų. Antarktikoje vabzdžiaėdžiai augalai augo prieš 6 milijonus metų. Kur jie dabar, ir kaip jie galėjo evoliucionuoti?
Taip pat buvo ledynmečių ir kitokių klimato pasikeitimų. Šiuos globalius pasikeitimus dalinai įtakojo atstumo nuo Saulės iki Žemės pokyčiai, Žemės pasvirimo kampas, žemynų tarpusavio padėties pasikeitimas. Tik prieš 15 tūkstančių metų nemažą Šiaurės Amerikos, Europos ir Azijos dalį, kurioje dabar yra ištisi vabzdžiaėdžių augalų arealai, gaubė kilometro storio ledo sluoksnis. Kur buvo šie augalai ledynmečio periodu? (daugiau žr. Degreef, 1989a, PDF, Decoding Diversity: Glacial Stragglers)

Aldrovanda vesiculosa
Kad dar labiau sutirštinti spalvas, reiktų paminėti visą eilę masinių išnykimų. Kreidos-terciaro periode prieš 65,5 milijonų metų išnyko gerokai daugiau nei dinozaurai. Labiausiai buvo paveikta Šiaurės Amerika, kur neišgyveno 50% augalų rūšių. Kas žino, kiek vabzdžiaėdžių augalų rūšių (o gal ir genčių) nustojo egzistuoti šiuo laikotarpiu. Kitas, mažiau globalus masinis išnykimas buvo prieš 15,5 milijonų metų. Labiausiai jis palietė šiaurinę Europą. Visi tokie faktoriai įtakoja gyvybės raidą mūsų planetoje. Jie taip pat apsunkina mūsų detalų supratimą apie tai, kaip vyko gyvybės evoliucija – labai jau daug trūkstamų mozaikos dalių.
Padėti sužinoti keletą datų gvazdikiečių kladogramoje gali du CPN (Carnivorous Plants Nesletter) žurnale spausdinti apžvalginiai straipsniai apie Aldrovanda iškasenas.
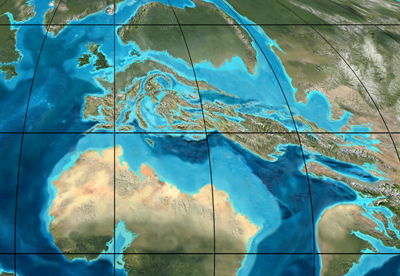
Satelitinis Europos vaizdas Eoceno periodu, prieš 50 milijonų metų. Šiuo laikotarpiu pirmosios aldrūnės (Aldrovanda) sėklos pavirto akmeniu salose, kurios vėliau tapo Europa © Ron Blakey, Northern Arizona University Geology.
Degreef (1997, PDF) apžvelgė publikacijas apie iškasenose rastas Aldrovanda sėklas ir žiedadulkes. (Iškasenos žinomos kaip Paleoaldrovanda splendens parodytos kaip vabzdžių kiaušiniai (Hermanova ir Kvacek, 2010). Šią dalį reiktų praleisti.) Seniausios pačios aldrovandos liekanos buvo rastos Europoje Eoceno laikotarpio uolienose, joms nuo 38 iki 55 milijonų metų. Atrasta įrodymų, kad nuo tada iki šių laikų išnyko 4 rūšys ir mažiausiai 6 gentys tapo nykstančiomis. Schlauer (1997, PDF) atkreipia dėmesį į 6 milijonų metų amžiaus iškaseną, kuri pasirodė esantis Aldrovanda lapas. Mes nežinome, kaip dar anksčiau atrodė Aldrovanda. Kadangi sėklos atrastos vandens nuosėdose, galima spėti, kad šie augalai visą laiką augo vandenyje ar pusiau panirę. (visą iškasenų duomenų santrauką žr. Cross, 2012)
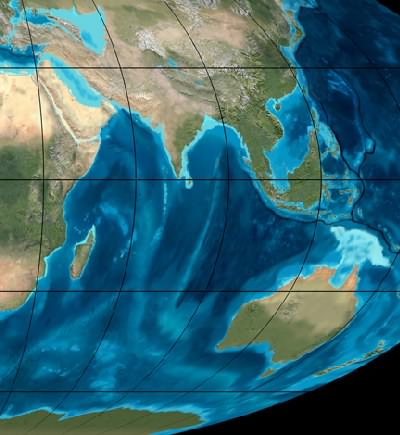
Indijos vandenyno satelitinis vaizdas mioceno laikotarpyje, prieš 20 milijonų metų. Šiuo laikotarpiu dėl klimato kaitos ir žemynų judėjimo migravo daugelis rūšių. Atkreipkite dėmesį, kad Antarktidos pakrantėje nebuvo ledo © Ron Blakey, Northern Arizona University Geology.
Iškasenų pavyzdžiai įrodo, kad Nepenthes ne visada augo ten, kur sutinkami dabar. Seniausios Nepenthes žiedadulkės rastos Eoceno nuosėdose, susidariusiose Tetijos vandenyno šiaurėje ir dabar esančios Europoje (Krutzsh, 1985). Šiuos duomenis patvirtino ir jų interpretaciją išplėtė Meimgerg ir Heubl (2006) DNR tyrimai, kuriais jie atrado, kad vakarų Indijos vandenyno populiacijos buvo sudarytos iš visiškai pažengusių Nepenthes rūšių dar prieš atsirandant (ar lygiagrečiai) Pietryčių Azijos populiacijoms. Buvo daugiau nei viena migracija į Pietryčių Aziją iš centrinės Azijos dalies, o taip pat, kintant vandenynų lygiui, buvo daugybė migracijų tarp salų. Tai reiškia, kad Miocene egzistavo visiškai išsivystę ąsoteniai.
Skirtingai nei Aldrovanda ir Nepenthes, seniausios žiedadulkių, panašių į saulašarių (Drosera) žiedadulkes, iškasenos rastos Australijoje ir Antarktikoje. Truswell ir Marchant (1986) aprašė žiedadulkes iš centrinės Australijos dalies, pavadindami augalus Fischeripollis halensis. Iškasenos, rastos netoli Elis Springso, aptiktos uolienose, susidariusiose ankstyvojo-viduriniojo Eoceno laikotarpiu. Macphail ir Truswell (2004) pranešė apie žiedadulkes, pavadintas Fischeripollis sp. A. Jos buvo rastos nuosėdiniuose sluoksniuose, išgręžtuose Antarktidos pakrantėje. Kai šių straipsnių nuotraukos buvo sulygintos su Takahashi ir Sohma (1982) nuotraukomis, paaiškėjo, kad Fischeripollis yra Drosera, o Fischeripollis sp. A labiau panašus į bendrą protėvį nei Fischeripollis halensis. Šios iškasenos byloja, kad saulašarės, tokios, kokias mes jas žinome, augo Australijoje ir Antarktidoje dar prieš 40 milijonų metų. Turint omeny, kiek nedaug saulašarių žieduose būna žiedadulkių, tai, kad jos buvo atrastos yra tiesiog stebuklas. Absoliuti dauguma atrastų iškasenose žiedadulkių buvo iš vėjo apdulkinamų žiedų. Antarktidos gręžiniuose su Fischeripollis žiedadulkėmis maždaug trečdalį sudarė panašios į notofagų (Nothophagus) žiedadulkės.
Iškasenų pavyzdžiai įrodo, kad Caryophyllales eilės gentys ir bent keletas pogenčių egzistavo Eoceno laikotarpiu, prieš 34 – 54 milijonus metų. Aldrovanda ir Nepenthes aptikti Šiauriniame pusrutulyje, o Drosera – Pietiniame. Tai nusako ir galimo bendro protėvio egzistavimo laikotarpį. Tai – vėlyvasis Kreidos periodas, prieš 65-90 milijonų metų, kadangi reikėtų mažiausiai 15 milijonų metų, kad augalų grupės išsiskirtų taip radikaliai. Įdomu, kad bent dvi gvazdikiečių vabzdžiaėdžių rūšys pergyveno Keidos periodo asteroido kataklizmą, įvykusį prieš 65 milijonus metų. Šios rūšys galėjo duoti pradžią bent jau dviems pagrindinėms evoliucijos linijom. Deja, apie šių linijų išsiskyrimo datas nieko nėra žinoma. Droseraceae ir Nepenthaceae žiedadulkės yra labai išskirtinės, ir ateityje galėtų būti atrastos senesnėse uolienose.
Prieš pradedant paskutiniojo bendrojo gvazdikiečių vabzdžiaėdžių protėvio aptarimą, reiktų žvilgtelėti į Afrikos vabzdžiaėdį Triphyophyllum peltatum. Šis piešinys iš laiško, 1972 metais atspausdinto CPN (PDF), pasako viską:
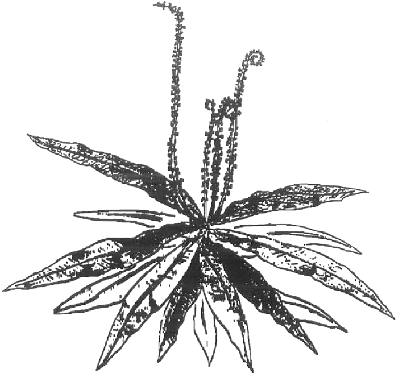
Jaunas augalas panašus į Nepenthes be ąsočių, su keletu Drosophyllum lapų, tik be žiedų jų galuose. Vabzdžiaėdžiai lapai su jautriaisiais plaukeliais, jie net gi išsivynioja taip pat, kaip tai daro Drosophyllum (atvirkštine spirale), visiškai priešingai, nei tai daro Drosera lapai. Drosera virškinamasis paviršius ir dauguma jautriųjų plaukelių yra vidinėje lapo pusėje, tuo tarpu Triphyophyllum ir Drosophyllum pirminis virškinamasis paviršius yra lapo išorėje. Lapai išsivynioja taip, kad pirminis virškinamasis paviršius atsiduria vidinėje skrotelės pusėje.

Triphyophyllum peltatum su trijų tipų lapais Bonos botanikos sodo šiltnamyje. Katja Rembold nuotrauka
Vabzdžiaėdžiai Triphyophyllum lapai turi lapkotį ir todėl atrodo kaip sudėtinių Nepenthes lapų analogai. Skirtumas tik tas, kad Nepenthes lapai iš tikrųjų yra sudėtiniai. Proto-Nepenthes pirminis virškinamasis paviršius buvo viršutinėje lapo pusėje, skirtingai nuo Triphyophyllum ir Drosophyllum.
Subrendę Triphyophyllum yra lianos. Lapai, išaugantys ant lianos stiebo, viršūnėse turi kabliukus, o Nepenthes šiose vietoje turi ąsočius. Lianos stiebai taip užtikrina geresnę augalo mitybą, kai vabzdžiaėdžiai lapai yra virš kabliukus turinčių lapų. Daugiau apie šį neįtikėtiną augalą galite pasiskaityti anglų kalba: Bringmann et al. (1999 PDF, 2002 PDF) ir McPherson (2008).
Būta daug diskusijų, ar paskutinysis bendras vabzdžiaėdžių gvazdikiečių protėvis buvo dalinai prisitaikęs ar pilnavertis vabzdžiaėdis augalas. Kitais žodžiais tariant, ar vabzdžiaėdiškumas šioje eilėje išsivystė vieną, du, tris ar daugiau kartų? Vienareikšmiškai atsakyti į šį klausimą neįmanoma. Kadangi augalas augo prieš 60 milijonų metų, reikia padaryti daug išvadų iš to, ką mes matom dabar. Ką mes tikrai žinome, tai, kad artimiausi vabzdžiaėdžių gvazdikiečių giminaičiai auga nederlingose augimvietėse ir daugelis turi liaukas, išskiriančias druskas arba gleives.
Ar galima pamatyti vabzdžiaėdiškumo ištakas vabzdžiaėdžiuose gvazdikiečiuose? Jie neatsirado tiesiog iš oro. Nepenthes fanai gali pagalvoti, kad atpažįsta augalą šiame Wilson (1890) piešinyje.
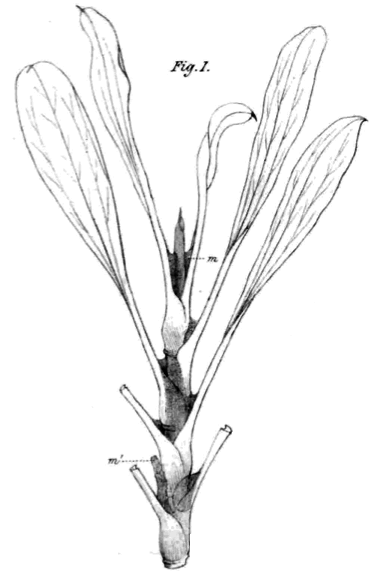
Jis atrodo panašus į ąsotenį be ąsočių, pavyzdžiui, Nepenthes rafflesiana. Deja, taip nėra. Nupieštas yra Limonium peregrinum (jūros gvazdikas, pelkių rozmarinas, jūros levanda), augantis Pietų Afrikos pakrantėse. Rūšis priklauso Plumbaginaceae šeimai, bet turi daug lapų ir liaukų požymių, būdingų tiek pliumbaginiams, tiek vabzdžiaėdžiams gvazdikiečiams. Augalo lapo pamatas išsiplėtęs ir apgaubiantis stiebą, o kai kurių kitų rūšių labai išvešėjęs, galintis atrodyti kaip ąsotis. Ši rūšis turi druskas išskiriančias liaukas lapkočio pamate. Juoda medžiaga, pavaizduota piešinyje varvanti stiebu, yra ne kas kita kaip gleivėtas skystis, kurį išskiria šios liaukos. Šis skystis kaupiasi prie lapo pamato ir gali nesunkiai užkirsti kelią stiebu ropojantiems vabzdžiams. Šis augalas ir jo giminaičiai turi daug charakteristikų, kurias, spėjama, turėtų turėti ir paskutinysis bendras vabzdžiaėdžių protėvis. Bet mes negalime tiksliai žinoti, kokie augalai buvo prieš 90 milijonų metų.

Nepenthes ventricosa
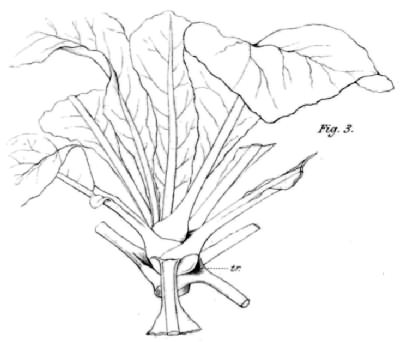
Limonium fruticans iš Kanarų salų. John Wilson (1890) piešinys. Išplėstuose prielapių pamatuose taip pat išsiskiria lipnus skystis
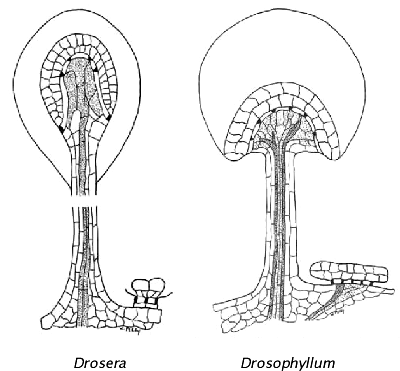
Paveikslėlis iš Haberlandt (1982 PDF), piešė Wayne Perry. Skirtinga jautriųjų plaukelių ir nejudrių liaukų struktūra. Drosophyllum jautrieji plaukeliai yra vaskuliarizuoti greitam skysčio išskyrimui, to nesimato Drosera jautriuose plaukeliuose.
Gvazdikiečių eilėje svarbiausios vabzdžiaėdžių liaukos yra nejudrios virškinamosios liaukos (žr. paveikslėlius dešinėje). Auką gaudantys Triphyophyllum ir Drosophyllum čiuptuvėliai išsivystė vėliau, o Drosera čiuptuvėliai yra daugiafunkciniai dariniai, jie perėmė nejudrių virškinimo liaukų funkcijas. Bendrumo tarp visų genčių virškinamųjų liaukų nėra, todėl galima manyti, kad šios liaukos evoliucionavo skirtingai, tačiau tai nėra įrodyta.
Drosera, Dionaea ir Aldrovanda linijų augalai yra arba vandens, arba pelkių augalai, tuo tarpu Nepenthes ir Drosophyllum linijose augalai prisitaikė prie gerai drenuotų augimviečių. Kai kurie autoriai, vadovaudamiesi ontogeneze ir fiziologija, teigia, kad Aldrovanda, Dionaea ir Drosera išsivystė iš vandeninio ar pusiau panirusio protėvio (Degreef, 1988, PDF), tačiau tokių įrodymų Nepenthes ar Drosophyllum linijose nėra. Paprasčiau yra įsivaizduoti, kad bendrasis protėvis buvo labiau panašus į Nepenthes ar Drosophyllum linijų augalus, nes pastarieji yra labiau panašūs į tipinius augalus, turinčius „normalias” šaknis ir lapus.
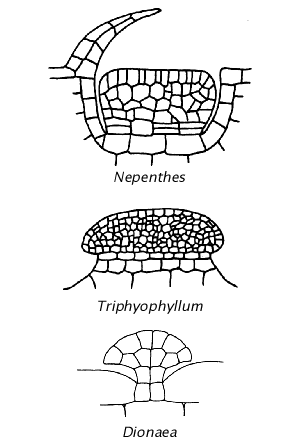
Müller (2002 PDF) virškinamųjų liaukų piešinys remiantis kitais šaltiniais. Nepenthes ir Triphyophyllum virškinamosios liaukos yra panašios į Drosophyllum, bet nėra vazkuliarizuotos. Nepenthes ąsotyje šios liaukos yra vidiniame paviršiuje ir išsidėsto vertikaliai, dengiamos ląstelių sluoksnio, neleidžiančio vabzdžiui ant jų užlipti. Triphyophyllum tipo liaukos sudaro tarsi kopėčias. Dionaea virškinammosios liaukos atrodo linkusios evoliucionuoti į jautrųjų plaukelį, bet nėra vazkuliarizuotos kaip saulašarių.
O kaip dėl bendro protėvio tarp Drosera ir Dionaea bei Aldrovanda linijų? Degreef (1988, PDF) šia tema pateikė neblogų samprotavimų. Įtikinamiausiai atrodo, kad bendras protėvis turėjo būti šlapynių ar pusiau paniręs vandens augalas. Remiantis šiandien išlikusių rūšių charakteristikomis, kaip streso atveju sutrinka jų lapų formavimasis bei lapų liaukų ir kitų darinių tipais, šis augalas turėjo būti panašus į Drosera arcturi (daugiau informacijos apie šią rūšį žr. Gibson, 1999 PDF). Protėvis galbūt neturėjo čiuptuvėlių, bet turėjau jautrias virškinamąsias liaukas lapo paviršiuje. Gali būti, kad lapai buvo judrūs ir pajutę auką, galėjo lėtai užsiverti. Primityvesnės veislės turi specifinius lapų pamato praplatėjimus, bet daugumos rūšių, įskaitant ir Dionaea bei Aldrovanda, lapai regresavo į pleištiškas ar šaukšto pavidalo formas, visų jų lapai turi nejudrias virškinamąsias liaukas, visų lapai judrūs. Visos gentys turi kitokias liaukas, kurių struktūra kiekvienoje gentyje skiriasi.
Yra ne viena hipotezė, nupasakojanti, kaip Dionaea galėjo evoliucionuoti iš sąlyginai modernios Drosera ar kito augalo su jautriaisiais plaukeliais. Visose šiose ganėtinai įtikinamose istorijose yra ekstremalių įvykių ir vystymosi pokyčių. Degreef (1988 PDF), remdamasis kitomis mokslinėmis publikacijomis, įvardija visą eilę priežasčių. Iš esmės, analoginės Dionaea ir Drosera struktūros yra bendro protėvio ir panašių evoliucijos kelių rezultatas, dėl ko išsivystė struktūros, kurios yra panašios, bet labai skiriasi jų detalės. Didžiausia evoliucijos prieštara – tu negali to patikrinti. Vabzdžiaėdžiai augalai tapo vabzdžiaėdžiai todėl, kad tai buvo vienintelis būdas jiems išgyventi. Tam, kad iš Drosera, kuri gaudo smulkučius vabzdžius, evoliucionuoti iki Dionaea, gaudančio stambesnes muses, reikia praeiti keletą tarpinių grandžių, kurių medžioklės efektyvumas gerokai mažesnis. Todėl evoliuciškai įtikinamiausia yra schema, kad abi gentys Drosera ir Dionaea išsivystė lygiagrečiai, veikiamo skirtingų aplinkos sąlygų.
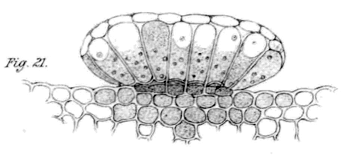
Limonium peregrinum druskas išskiriančios liaukos. Wilson (1890) piešinys
Dionaea ir Aldrovanda išvystė savo gaudykles atskirai. Jeigu pažiūrėsite, kaip veikia Dionaea ir Aldrovanda gaudyklės, suprasite, kad Aldrovanda nėra vandeninė Dionaea versija. Aldrovanda gaudyklės veikia visai kitu principu. Poppinga ir Joyeux (2011) pademonstravo, kaip musėkauto gaudyklės užsiveria išsiplečiant išoriniam jų ląstelių sluoksniui, dėl ko gaudyklės dalys išlinksta ir ji užsiveria. Tuo tarpu aldrūnės gaudyklės dalys susiglaudžia per susijungimą, pačios formos nekeisdamos. Reiškia, abiejų augalų gaudyklės išsivystė nepriklausomai. Šis faktas nebuvo žinomas, kai Degreef pateikė apžvalgą CPN.

Dionaea muscipula

Dionaea muscipula gaudyklės
Kaip išsivystė trijų dalių Nepenthes lapai? Suprasti mums gali padėti juveniliniai ąsotenių sėjinukų lapai. Kai kurie autoriai teigia, kad Nepenthes ąsočiai susiformavo plaukelių galiukų. Tai vienareikšmiškai klaidinga nuomonė.
Suaugę Nepenthes lapai turi tris dalis: platų žalią prielapį lapo pamatinėje dalyje, ūselį vidurinėje dalyje ir ąsotį lapo gale. Jaunų sėjinukų prielapis yra labai trumpas, o ūselio išvis nėra. Ąsočiai gali būti lape ar atskirai, išaugę iš pat lapo. Sėjinukams augant, priklausomai nuo rūšies, ąsotėlis išauga tiesiog iš prielapio. Dar vyresniuose augaluose pradeda formuotis ūselis tarp prielapio ir ąsotėlio.

Juveniliniai Nepenthes rafflesiana lapai. Kairėje esančio jaunesnio augalo lapo prielapis tiesiogiai pereina į ąsotėlį, o vyresnio augalo dešinėje tarp prielapio ir ąsotėlio pradeda formuotis stiebelis.
Dabartinė trijų dalių lapo struktūra išsivystė tam, kad ąsotėliai galėtų būti nutolę kuo didesniu atstumu nuo augalo skrotelės, arba kad augalui augant liana galėtų užsikabinti už atramų. Platus prielapis užtikrina efektyvią fotosintezę, o ilgi ūseliai užtikrina augalo užsikabinimą už atramų. Owen ir Lennon (1999) parašė išsamų straipsnį apie tai, kaip vystosi suaugę ąsočiai ir kokia jų liaukų struktūra.
Kaip ir kitoje šeimos medžio dalyje, taip ir čia – suprasti Nepenthes ir Drosophyllum adaptacijų evoliuciją yra sunku. Iš esmės nėra įmanoma, kad Triphyophyllum ir Drosophyllum išsivystė iš augalo, kurį mes vadiname Nepenthes. Nepenthes augalai yra dvinamiai – kiekvienas augalas turi arba moteriškus, arba vyriškus žiedus. Tuo tarpų Triphyophyllum ir Drosophyllum augalų žiedai hermafroditiniai (abiejų lyčių). Įmanoma, kad dvinamis augalas evoliucijos eigoje gali išsivystyti taip, kad turėtų ir moteriškus, ir vyriškus žiedus, tačiau tam, kad susijungtų abi lytis viename žiede, turi įvykti kažkas unikalaus. Be to, Nepenthes rūšys yra oktaploidinės, turinčios 8 chromosomų poras, tuo tarpu Triphyophyllum yra tetraploidas, o Drosophyllum – diploidas.

Nepenthes aristolochioides ąsotis
Taip pat neįtikėtina, kad kažkas panašaus į Triphyophyllum ar Drosophyllum išsivystytų iki panašaus į Nepenthes. Abu lipnialapiai augalai yra labai specializuoti, o jų pirminis virškinamasis paviršius yra kitoje nei Nepenthes lapo pusėje. Paprasčiausias paaiškinimas būtų, kad rūšys, turinčios virškinamuosius paviršius abiejuose lapų pusėse išsiskyrė į dvi rūšis, kurios vystėsi skirtingomis kryptimis. Vienos linijos augalai vystėsi stengdamiesi apsaugot savo virškinamąjį lapo paviršių nuo lietaus poveikio, kiti – vystė jautriuosius plaukelius išorinėje lapo pusėje.
Žiupsnelis druskos DNR tyrimams
Lyginant rūšis genties viduje, DNR tyrimais galima išsamiai paaiškinti ryšius tarp esamų rūšių ir pateikti bendrus patarimus geno paleogeografijai. Tačiau net jei visos šiandien egzistuojančios rūšys būtų įtrauktos į genų analizę, vis tiek atsirastų didžiulių spragų techniškai sudarant giminystės medžius. Viena iš pagrindinių priežasčių – pagrindinių rūšių neįtraukimas į duomenų analizę. Todėl kladogramos gali parodyti menamas asociacijas ir akivaizdžius ryšius, kurių realiai niekada nebuvo. Visi DNR tyrimų rezultatai turi būti siejami su kitais fiziniais duomenimis tam, kad įsitikinti, ar jie turi prasmės.
Peržvelgdami DNR duomenis, mes turime priimti domėn, kad buvo begalės tarprūšinių kryžminimųsi Nepenthes ir Drosera gentyse. DNR sekos analizė priima nepriklausomą rūšių linijų evoliuciją. Tarprūšinis kryžminimasis šiuos duomenis iškraipo. Daugelis Nepenthes rūšių kryžminasi tarpusavyje, dauguma Drosera rūšių gali turėti hibridus. Hibridizacijos pasekmė – mišrūs palikuonys. Jei analizuojama tik dalis DNR sekos ar tik plastidžių DNR seka, gali pasirodyti, kad analizuojame vieną iš tėvinių rūšių. Pridėjus daugiau sekų (ypač iš branduolio DNR), gali paaiškėti, kad turime neegzistuojančią liniją. Jei tyrimai apima ir branduolio, ir plastidžių sekas, jų rezultate gauti giminystės medžiai gali būti iškreipti, lyginant su fragmentiniais tyrimais.
Mes turime sumažinti rūšių skaičių, kadangi dauguma Australijos saulašarių rūšių turi vidurūšinį nesuderinamumą, o kai kurios ąsotenių rūšys turi tik vyriškus arba tik moteriškus augalus. Tam, kad ąsoteniai užmegztų sėklas, reikia abiejų – vyriško ir moteriško – individų, o tam, kad apsidulkintų vidurūšinį nesuderinamumą turinčios saulašarės, reikia žiedadulkių, kurios genetiškai pakankamai tolimos motininiam augalui. Tokie „nutolę” augalai gali būti kitos rūšys. Būtent todėl yra daugybė endeminių rūšių. Šie augalai pasmerkti neužmegzti sėklų ir neišplisti toliau. Situacija gali būti dar įdomesnė, jei vienos rūšies augalai atsiranda kitos rūšies teritorijoje. Patys šie augalai negali užmegzti sėklų, bet gali apdulkinti seniau buvusių augalų žiedus – taip gali susidaryti naujos rūšys.
Plačiai paplitusios Australijoje bei migravusios į Naująją Zelandiją bei kitus kontinentus, saulašarių rūšys turi vidurūšinį suderinamumą. Jų paplitimui užtenka, kad bent viena sėkla pasiektų kitą saulašarių populiaciją. Šis reiškinys buvo atrastas DNR sekos analizės pagalba.

Drosera regia lapas. Dideli jautrūs plaukeliai ir sėslios mažos raudonos virškinamosios liaukos pačiame lapo paviršiuje
Rivadavia et al. (2003) atliko išsamią saulašarių DNR studiją. Mano manymu, ji nebuvo labai išsami, nes nebuvo įtrauktos kai kurios svarbios rūšys. Nepaisant to, studija pateikė labai įdomius rezultatus. Išnagrinėjęs studiją 2010 metais ir pasinaudojęs visais tuo metu viešai prieinamais duomenimis bei Rivadavia et al. (2003) duomenimis, aš pateikiau apibendrinimą Dorsera filogenezės puslapyje.
Remiantis DNR duomenimis, seniausios vis dar egzistuojančios rūšys yra Afrikoje (Drosera regia) ir Australijoje bei Naujojoje Zelandijoje (Drosera arcturi). Ne visos galimos senovinės rūšys buvo tyrinėtos, taigi, galimas atvejis, kad jų yra ir daugiau, ypač Pietų Amerikoje. Tai patvirtintų ir žiedadulkių duomenis bei nurodytų pietų pusrutulio židinį. Kreidos periode pietiniai žemynai buvo gerokai arčiau vienas kito, nebuvo Vakarų vėjų srovės, klimatas buvo šiltesnis, todėl galėjo būti daugiau gyvybės judėjimo tarp žemynų. Tačiau nereikia atmesti ir vėlyvesnio Drosera regia migravimo dideliu atstumu. Visais atvejais Drosera regia dabar nerandama Afrikos floroje. Drosera arcturi auga sub-alpiniuose Australijos, Tasmanijos ir Naujosios Zelandijos regionuose, ir su anksčiau minėta rūšimi neturi jokių artimų ryšių.

Drosera regia iš Pietų Afrikos. Augalai apie 40 cm aukščio. Drosera regia labai skiriasi nuo kitų saulašarių rūšių, todėl 1996 metais buvo siūloma jos pavadinimą pakeisti į Freatulina regia. Pavadinimo keitimui nebuvo pritarta.
Australijoje labai daug Drosera rūšių, kurių dauguma yra labai siauros specializacijos. Nė viena iš pūkuotųjų, gumbinių, stiebinių, nykštukinių saulašarių nėra aptinkama kituose žemynuose. Tokia didelė endeminių rūšių įvairovė rodo, kad jos turi genetini vidurūšinį nesuderinamumą. Drosera glanduligera ir Drosera burmannii turi vidurūšinį suderinamumą, yra vienmetės, tačiau jos neevoliucionavo į dešimtis skirtingų rūšių.
Iš Rivadavia duomenų aišku, kad viena ar keletas sąlyginai primityvių Drosera rūšių migravo iš Australijos ar Antarktidos į Šiaurės pusrutulį pro Aziją. Skirtingi migrantai nukeliavo į Pietų Ameriką ir Afriką galimai mioceno periodu ar vėliau. Šiaurės ir Pietų Amerikos linijos saulašarės galėjo migruoti tarp šių žemynų. Rivadavia tyrimuose nėra rūšies, kolonizavusios visą pasaulį. Tačiau vėlesni tyrimai parodo, kad Drosera spatulata yra stipri kandidatūra į bazinės rūšies daugeliui Amerikos, Afrikos ir Azijos rūšių poziciją. Šie tyrimai taip pat parodo, kad galimai Drosera spatulata yra ne viena rūšis, o rūšių grupė. Kad geriau suprasti šią rūšį bei rūšių judėjimą iš Australijos ir/arba Antarktidos, reikia atlikti daugiau DNR tyrimų.

Australijos Drosera peltata skrotelės fazė
Afrikos rūšys (neskaitant Drosera regia) yra glaudžiai susijusios tarpusavyje, tačiau Rivadavia analizė negalėjo nustatyti jų giminystės medžio. Tai gali būti spindulinės invazijos į žemyną ar hibridizacijos rezultatas, arba abi priežastys.
Yra žinomi tik keli tarpžemyniniai šuoliai, sukūrę giminingas rūšis. Drosera sessilifolia rytinėje Pietų Amerikoje pasirodė labai artima ir atsiskyrusi nuo Australietiškos ir pietų Azijos Drosera burmannii. Drosera uniflora iš pietrytinės Pietų Amerikos dalies atsiskyrė nuo Naujosios Zelandijos Drosera stenopetala. Yra ir daugiau Pietų Amerikos rūšių, atsiskyrusių nuo dideliu atstumu nuo savo giminaičių. Vienas pavyzdys yra Drosera meristocaulis, auganti šiaurės rytinėje Pietų Amerikos dalyje. Tai primityvi nykštukinė saulašarė, artima Drosera scorpioides, augančiai Australijoje (Rivadavia et al. 2012).

Juveniliniai Drosera binata iš Tasmanijos lapai. Suaugęs augalas turi šakutės pavidalo lapus, dėl kurių šis augalas ir gavo savo pavadinimą.
Daugiau informacijos galima rasti šiuose šaltiniuose, tačiau būtina atkreipti dėmesį, kad kiekvienas autorius pateikia savo turimais duomenimis suformuotą nuomonę, kurią nesunkiai gali pakeisti naujesni atradimai.
- Gerhard Bringmann, Jan Schlauer, Kristina Wolf, Heiko Rischer, Uwe Buschbom, Andreas Kreiner, Friedrich Thiele, Martin Duschek, Laurent Ake Assi, 1999, Cultivation of Triphyophyllum peltatum (Dioncophyllaceae), the part-time carnivorous plant, Carnivorous Plants Newsletter 28(1):7-13 (PDF),
- Bringmann, Gerhard and Heiko Rischer and Jan Schlauer and Kristina Wolf and Andreas Kreiner and Martin Duschek and Laurent Ake Assi, 2002, The Tropical Liana Triphyophyllum peltatum (Dioncophyllaceae): Formation of Carnivorous Organs is only a facultative prerequisite for shoot elongation, Carnivorous Plants Newsletter 31(2):44-52 (PDF),
- Kenneth Cameron, Kenneth J. Wurdack, Richard W. Jobson, 2002, Molecular evidence for the common origin of snap-traps among carnivorous plants, American Journal of Botany 89(9) 1503-1509,
-

Drosera spatulata iš Naujosios Zelandijos pietų. Augalas yra apie 20 mm skersmens.
A. Cross, 2012, Aldrovanda the Waterwheel Plant, Redfern Natural History Productions, Poole, Dorset, England,
- John D. Degreef, 1988, The Evolution of Aldrovanda and Dionaea Traps, Carnivorous Plants Newsletter 17(4):119-125 (PDF),
- John D. Degreef, 1989a, The Droseraceae during the glaciations, Carnivorous Plants Newsletter 18(2):45-46, 52-54 (PDF),
- John D. Degreef, 1989b, Early history Drosera and Drosophyllum, Carnivorous Plants Newsletter 18(3):86-89 (PDF),
- John Degreef, 1990a, Evolutionary patterns in Drosera, Carnivorous Plants Newsletter 19(1-2):11-16 (PDF),
- John D. DeGreef, 1990b, More on the evolution of Drosera, Carnivorous Plants Newsletter 19(3-4):92 (PDF),
- John D. Degreef, 1997, Fossil Aldrovanda, Carnivorous Plants Newsletter 26(3):93-97 (PDF),
-

Drosera spatulata iš Honkongo. Augalas apie 35 mm skersmens.
Robert Gibson, 1999, Drosera arcturi in Tasmania and a comparison with Drosera regia, Carnivorous Plants Newsletter (3):76-80 (PDF),
- Gottlieb Haberlandt, (iš Sinnesorgane Im Pflanzenreich, bert Carla R. Powell), 1982, Insectivores: Drosera and Drosophyllum, Carnivorous Plants Newsletter 11(3):66-73 (PDF),
- Z. Hermanova, J. Kvacek, 2010, Late Cretaceous Palaeoaldrovanda, not seeds of a carnivorous plant, but eggs of an insect, Journal of the National Museum (Prague), Natural History Series, 179(9):105–118,
- G. Heubl, G. Bringmann, H. Meimberg, 2006, Molecular phylogeny and character evolution of carnivorous plant families in Caryophyllales – revisited, Plant Biology 8(2006):821-830,
- Jonathan, 1992, A letter from Sierra Leone, Carnivorous Plants Newsletter 21(3):51-53 (PDF),
-

Drosera tokaiensis iš Japonijos. Ši rūšis yra hibridinė. Tėvai yra Drosera spatulata ir Drosera rotundifolia.
W. Krutzsh, 1985, Uber Nepenthes-Pollen (alias „Droseridites” p.p.) im europaischen Tertiar, Gleditschia 12:89-93,
- W. Krutzsh, 1989, Paleogeography and historical phytogeography (paleochorology) in the Neophyticum, Plant Systematic and Evolution 162:5-61,
- M. Dolores Lledo, Manuel B. Crespo, Kenneth M. Cameron, Michael F. Fay, Mark W. Chase, 1998, Systematics of Plumbaginaceae Based upon Cladistic Analysis of rbcL Sequence Data, Systematic Botany 23(1):21-29,
- M.K. Macphail, E.M.Truswell, 2004, Palynology of Site 1166, Prydz Bay, East Antarctica. In Cooper, A.K., O’Brien, P.E., and Richter, C. (Eds.), Proc. ODP, Sci. Results, 188:1–43. (PDF),
- Stewart McPherson, 2008, Glistening Carnivores: the sticky-leaved insect-eating plants, Redfern Natural History Productions,
-

Drosera hirtella iš Pietų Amerikos
H. Meimberg, G. Heubl, 2006, Introduction of a Nuclear Marker for Phylogenetic Analysis of Nepenthaceae, Plant Biology 8(2006):831–840,
- K. Müller, Th. Borsch, L. Legendre, I Theisen, W. Barthlott, 2002, Evolution of carnivory in the Lentibulariaceae: considerations based on molecular, morphological, and physiological evidence, Proc. 4th Intl. Carniv. Pl. Conf. pages 63-73 (PDF),
- T. P. Owen, Jr. ir K. A. Lennon, 1999, Structure and development of the pitchers from the carnivorous plant Nepenthes alata (Nepenthaceae), American Journal of Botany 86(10):1382–1390 (PDF),
- Simon Poppinga, Marc Joyeux, 2011, Different mechanics of snap-trapping in the two closely related carnivorous plants Dionaea muscipula and Aldrovanda vesiculosa, Physical Review E 84, 041928. DOI: 10.1103,
-

Drosera dielsiana iš Pietų Afrikos
Katja Rembold, Andreas Irmer, Simon Poppinga, Heiko Rischer, Gerhard Bringmann, 2010, Propagation of Triphyophyllum peltatum (Dioncophyllaceae) and observations on its carnivory, Carnivorous Plants Newsletter 39(3):71-77,
- Fernando Rivadavia, Katsuhiko Kondo, Mitsuyasu Hasebe, 2002, Phylogeny of the sundews, Drosera (Droseraceae) based on chloroplast rbcL and nuclear 18S ribosomal DNA sequences, Proc. 4th Intl. Carniv. Pl. Conf. pages 9-13 (PDF) (incomplete),
- Fernando Rivadavia, Katsuhiko Kondo, Masahiro Kato, Mitsuyasu Hasebe, 2003, Phylogeny of the sundews, Drosera (Droseraceae) based on chloroplast rbcL and nuclear 18S ribosomal DNA sequences, American Journal of Botany 90(1):123-130,
- F. Rivadavia, V. F. O. de Miranda, G. Hoogenstrijd, F. Pinheiro, G. Heubl, A. Fleischmann, 2012, Is Drosera meristocaulis a pygmy sundew? Evidence of a long-distance dispersal between Western Australia and northern South America. Ann Bot 110(1): 11-21,
- Jan Schlauer, 1997, ‘New’ data relating to the evolution and phylogeny of some carnivorous plant families. Carnivorous Plants Newsletter 26(2):34-38,
- Jan Schlauer, 1997, Fossil Aldrovanda — Additions, Carnivorous Plants Newsletter 26(3):98 (PDF),
- A. Stoltzfus, J. Suda, R. Kettering, A. Wolfe, S. Williams, 2002, Secretion of digestive enzymes in Plumbago. Proc. 4th Intl. Carniv. Pl. Conf. pages 203-207 (PDF),
- H. Takahashi, K. Sohma, 1982, Pollen morphology of the Droseraceae and its related taxa. Sci. Rep. Tohoku Univ., IV 38(2):81-156,
- E.M. Truswell, N.G. Marchant, 1986, Early Tertiary pollen of probable Droseracean affinity from central Australia. Spec. Pap.—Palaeontol., 35:163–178,
- Jaohn Wilson, 1890, The mucilage- and other glands of the Plumbagineae. Ann. Bot. 4:231-258,
- C. Yesson, A. Culham, 2006, Phyloclimatic Modeling: Combining Phylogenetics and Bioclimatic Modeling, Systematic Biology 55(5):785-802.

Drosera brevifolia iš Šiaurės Amerikos. Šši švelnias žiemas mėgstanti saulašarė yra glaudžiai susijusi su šaltas žiemas ištveriančia ir žieminius pumpurus formuojančia Drosera rotundifolia

Drosera rotundifolia sutinkama visame Šiaurės pusrutulyje
.
.
.
.
.
.
.
.
.
.
John Brittnacher
Vertė J. Mackevičius
.
© International Carnivorous Plants Society (ICPS). Visa šiame straipsnyje esanti informacija kitose informacijos sklaidos priemonėse gali būti naudojama tik gavus raštišką ICPS sutikimą.